|
第三章
补体系统
补体(complement):存在于正常人及动物血清中的一组与免疫相关并具有酶活性和自我调节作用的球蛋白。这些蛋白是抗体发挥免疫溶菌、溶细胞作用的必要补充条件,故称补体。包括近40种可溶性蛋白、膜结合蛋白和补体受体蛋白,故又称补体系统(Complement
System)。
第一节
补体系统的组成
一、补体系统的组分和命名
根据功能可将补体系统成分分为五组:
1.参与经典活化途径的补体成分:
C1r、C1s、C1q、C4、C2。
2.参与旁路活化途径的补体成分:
B因子、 D因子、C3。
3.参与MBP活化途径的补体成分:
MBL、丝氨酸蛋白酶。
4.共同末端通路的成分:: C3、
C5、 C6、 C7、 C8和C9。
5.调控补体活化的蛋白分子及补体受体
可溶性调节分子:C1抑制物、H因子、I因子、C4bP、S蛋白等。
膜结合调节分子:膜辅助因子蛋白、同源限制因子、膜反应溶解抑制因子等。
补体受体:CR1,CR2,CR3/CR4及CR5a。
二、补体组分的理化特性
:
1.均为球蛋白,含量相对稳定。
含量最高的成分是
C3 ;含量最少的是 D 因子。
分子量最大的是
C1q ;分子量最小的是 D 因子。
2.以无活性的酶前体形式存在,被激活后才能发挥生物效应。
3.性质极不稳定,易受理化因素影响而灭活。例如:56℃30分钟灭活。
4.多种细胞均能合成补体:肝细胞和巨噬细胞是产生补体的主要细胞。
第二节
补体系统的激活与调节
补体系统的激活呈放大的连锁反应,前一组活化的成分是后一组催化酶。根据激活过程中起始顺序不同,可分为三条途径:即经典途径、旁路途径和MBL途径。
一、补体激活的经典途径
经典途径----由抗原抗体复合物激活补体的途径。
病原生物进入机体,能诱导免疫系统产生抗体,抗体产生后,可以与病原生物的抗原物质结合形成抗原抗体复合物,因此,经典途径的激活在抗体产生之后。
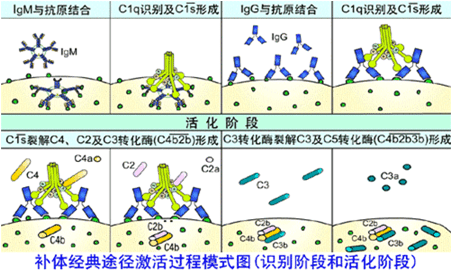
经典途径从C1q激活开始,继而相继激活C4、C2、C3,最终形成C5转化酶。
1.C1激活
抗原抗体形成复合物后构型改变,暴露补体结合位点供C1q识别并结合。
C1q必须同时与两个以上的活化位点结合才能被激活(1个IgM分子即可激活补体,2个以上IgG才能激活补体),C1q与抗体结合后导致C1r、C1s激活。
2.C3转化酶(C4b2b)形成
在Ca2+参与下,活化的C1s可以依次裂解C4和C2成为C4a、C4b和C2a、C2b,
然后C4b与C2b结合形成C4b2b,即C3转化酶形成。
3.C5转化酶(C4b2b3b)形成:
C3转化酶裂解C3为C3a和C3b,C3b与C4b2b结合形成C3b4b2b,即C5转化酶(C3b4b2b)形成。
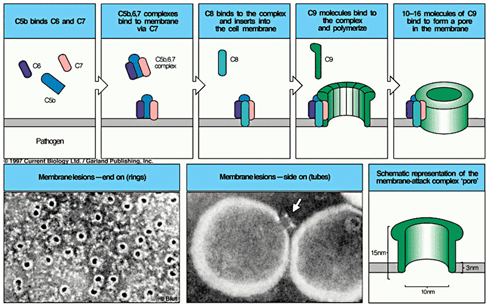
4.攻膜单位(C5b6789)形成:
C5转化酶裂解C5成为C5a和C5b开始,经C6、C7、C8、C9的相继激活,最终形成C5b6789大分子复合物即攻膜单位(membrane
attack complex,MAC),导致细胞溶解。
二、补体激活的替代途径
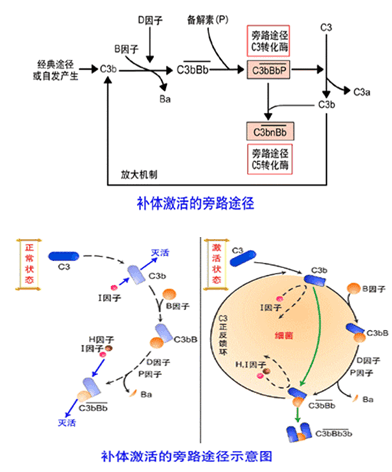
替代途径从C3激活(C3自发裂解)开始, B因子和D因子参与的激活过程,最终形成C5转化酶。
激活物质----细菌内毒素,酵母多糖,葡聚糖,凝聚的IgA、IgG4等。
1.C3激活:
体液中少数C3分子在生理情况下自发裂解为C3a和C3b。此时如有替代途径激活物质存在,C3b得以黏附于激活物的表面。黏附于激活物表面的C3b可以与B因子结合也可以与C5结合。
2.C3转化酶形成:
黏附于激活物表面的C3b与B因子结合为C3bB,在D因子作用下形成C3bBb,即替代途径的C3转化酶形成。
3.C5转化酶形成:
C3转化酶裂解C3为C3a和C3b。C3b与C3bBb结合成为C3bBb3b或C3bnBb,即替代途径的C5转化酶形成。
4.C3b正反馈环路:
血清中的P(propedin,备解素)与C3bBb结合使之稳定。有利于C3bBb能更多地裂解C3,形成C3b正反馈环路。
5.补体攻膜单位:
细胞膜表面的C3b5b与C6、C7、C8依次结合形成C5b678复和物。
该复合物诱发C9在细胞膜表面共聚,膜表面形成通道造成胞膜的穿孔损
三、补体激活的MBP途径
MBP途径是由甘露聚糖结合凝集素MBL与细菌甘露糖残基和丝氨酸蛋白酶结合启动激活补体的途径。在病原微生物感染的早期,体内巨噬细胞、中性粒细胞可产生TNF-α、IL-1和IL-6,从而导致机体发生急性期反应,并诱导肝细胞合成分泌急性期蛋白,其中参与补体激活的有MBL和C反应蛋白。
正常血清中MBL水平极低,在急性期反应时其水平明显升高。MBL与C1q分子结构类似。MBL首先与细菌的甘露糖残基结合,然后与丝氨酸蛋白酶结合,形成MBL相关的丝氨酸蛋白酶(MASP),MASP具有与活化的C1q同样的生物学活性,可水解C4和C2分子,继而形成C3转化酶,其后的反应过程与经典途径相同。C反应蛋白可与C1q结合并使之激活,然后依次激活补体其他成分。
四、补体活化过程的调节
1.补体的自身调控:C3转化酶极易衰变。
2.补体调节因子的作用:C1抑制物与活化的C1r、C1s及MASP结合,从而抑制C1的活化;I因子裂解游离的和结合状态的C3b和C4b,使C3转化酶(C4b2b)不能形成;
第三节
补体系统的生物学功能
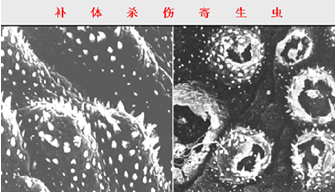
一、溶菌、杀菌及细胞毒作用:
革兰阴性细菌、有包膜病毒、红细胞、血小板及某些其它细胞可通过经典或旁路途径激活补体,由攻膜单位导致膜结构损伤而使细菌、细胞溶解。这种杀菌和溶解细胞作用可由特异性抗体协助完成,也可由于直接激活补体而产生。
二、调理作用:
免疫复合物激活补体形成的复合物中的C3b,能与吞噬细胞表面的C3b受体结合,通过C3b的桥联作用将抗原抗体补体复合物结合在吞噬细胞表面,促进了吞噬细胞对抗原的吞噬破坏,此称为依赖补体的调理吞噬作用。
三、免疫粘附与清楚免疫复合物:
抗原抗体复合物激活补体,可通过补体的C3b粘附于表面有C3b受体的红细胞、血小板上,形成较大的复合物,有利于吞噬细胞的吞噬清除,这对抗感染免疫有重要作用。
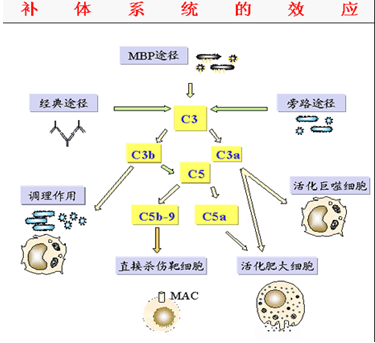
四、炎症介质作用
1.激肽样作用:游离的C2a具有激肽样作用,能引起局部炎
症充血和水肿。
2.过敏毒素作用:C3a、C5a可使肥大细胞、嗜碱性粒细胞释放组胺,引起局部
血管扩张,通透性升高,肠道平滑肌收缩和支气管痉挛。
3.趋化作用:C3a、C5a、C567具有趋化因子作用,能吸引吞噬细胞至感染部位发挥吞噬作用。
|